Célula espermática > Cauda ou flagelo > Saiba mais
A estrutura e função do axonema têm sido estudadas na alga verde unicelular Chlamydomonas reinhardtii. Nesta espécie, já foram identificadas mais de 200 proteínas do axonema e associadas a este.
Estrutura do axonema: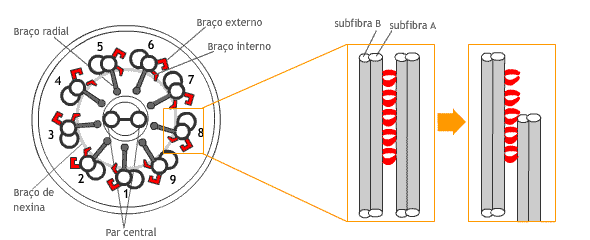
Alguns autores denominam os microtúbulos centrais de microtúbulo centrais 3 e 8 (MC 3 e 8), pois ficam próximos dos diplo microtúbulos 3 e 8, respectivamente. Do par central, partem braços radiais que se conectam aos 9 diplomicrotúbulos, formando uma hélice dupla ao redor do par central Os 9 diplo microtúbulos do axonema recebem numeração de 1 a 9. Cada um deles é formado de duas subfibras de α e ß tubulina e são denominadas fibras A e B. Além da tubulina, entretanto, inúmera outras proteínas provavelmente estejam também associadas a esta estrutura, tal como ocorre na Chlamydomonas.
 |
Subfibra A: é um microtúbulo completo (de forma cilíndrica), composto de 13 protofilamentos e sobre o qual está colocado uma subfibra B. Subfibra B: é mais curta, possui 10-11 protofilamentos. É um cilindro incompleto (em forma de “C”), o que permite que deslize ao longo do microtúbulo A adjacente. |
Do microtúbulo A, duas estruturas , denominadas braços – (braço interno e braço externo) são estendidas ao microtúbulo B do próximo par de microtúbulos.
Os braços são formados por dineína, proteínas que hidrolisam o ATP e são consideradas como o “motor” flagelar.
A ativação da dineína ATPase faz com que ocorra o deslizamento dos microtúbulos adjacentes, o que resulta na ondulação do flagelo. A movimentação flagelar ocorre pela atividade dos braços de dineína, trabalhando contra a estrutura estável dos diplomicrotúbulos.
Os diplomicrotúbulos são conectados entre si através de braços de nexina.
No início da espermiogênese, o flagelo espermático é visualizado projetando-se da superfície de uma espermátida arredondada. O axonema é a primeira estrutura do flagelo que pode ser observada e origina-se do centríolo distal (ou posterior). Este se localiza no pólo caudal do núcleo e alonga-se para uma direção distal.
Os centríolos funcionam como centros organizadores para a formação do axonema e das colunas segmentadas. O centríolo proximal é considerado como o centro da motilidade espermática, embora também existam espécies em que o flagelo apresenta motilidade normal e o centríolo proximal tenha desaparecido nos estágios finais da espermiogênese.
Em alguns cílios, a estrutura do axonema não apresenta o par central (a estrutura é 9+0) e são denominados de cílios primários. Mais recentemente, se considera que existam quatro tipos de cílios: cílios móveis (9+2), no trato respiratório, ductos eferentes e flagelo espermático; cílios móveis (9+0), no nodo embrionário, cílios sensoriais (9+2) vestibulares, cílios sensoriais (9+0) no rim e córnea. Alterações destas estruturas causam uma série de patologias em mamíferos.
Os cílios e flagelos possuem a mesma estrutura 9+2, mas os cílios são mais curtos, com menos de 10 µm de comprimento, enquanto que os flagelos são cílios mais longos, com até 200 µm.